
Original:http://www.tulane.edu/~wiser/protozoology/notes/mal_lc.html
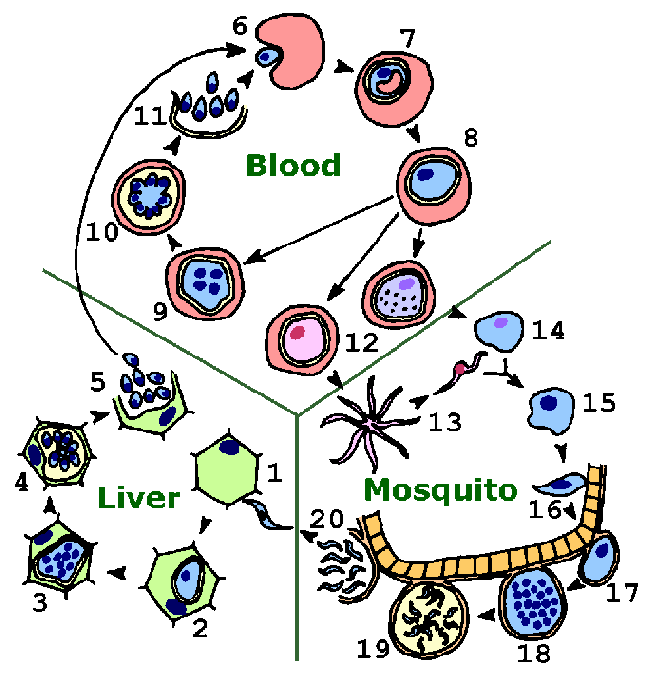
Малярийный паразит проявляет сложный жизненный цикл, включающий вектор насекомых (москит) и хозяина позвоночных (человека). (См. Также файл в формате pdf для печати.) Основными этапами жизненного цикла являются: стадия в печени, стадия в крови, половая стадия и спорогония.
История
Инфекция начинается, когда спорозоиты вводятся слюной кормящегося комара. Спорозоиты переносятся кровеносной системой в печень и вторгаются в гепатоциты (1). Внутриклеточный паразит подвергается бесполой репликации, известной как экзоэритроцитарная шизогония внутри гепатоцитов (2-4). Экзоэритроцитарная шизогония достигает высшей точки в производстве мерозоитов, которые высвобождаются в кровоток (5). Часть паразитов печени-стадии от P. vivax и P. ovale проходят через неактивный период (не показан) вместо того, чтобы немедленно пройти через бесполошную репликацию (то есть временно остановиться на этапе 2). Эти гипнозоиты активируются через несколько недель или месяцев (или лет) после первичной инфекции и несут ответственность за рецидивы. (Возврат к рецидивам в основном документе.)
Мерозоиты вторгаются в эритроциты (6) и подвергаются трофическому периоду, в течение которого паразит увеличивается (7-8). Ранний трофозоит часто называют «кольцевой формой» из-за его морфологии. Укрупнение трофозоитов сопровождается активным метаболизмом, включая поглощение цитоплазмы хозяина и протеолиза гемоглобина в аминокислоты. Окончание трофического периода проявляется в многократных раундах ядерного деления без цитокинеза, в результате получается шизонт (9). Мерозоиты растут из зрелого шизонта, также называемого сегментирующим (10), и мерозоиты высвобождаются после разрыва инфицированного эритроцита (11). Вторжение эритроцитов возобновляет очередной цикл репликативного цикла крови (6-11). (Вернитесь к шизогонии в главном документе.)
Прерывистые лихорадки, часто связанные с малярией, вызваны синхронным разрывом инфицированных эритроцитов и выделением мерозоитов (см. Малярийный пароксизм). Трофозоит- и шизонт-инфицированные эритроциты редко обнаруживаются в периферическом кровообращении при инфекциях P. falciparum. Эритроциты, инфицированные этими стадиями, прилипают к эндотелиальным клеткам и секвестрируют микрососуды жизненно важных органов, особенно мозга, сердца и легких. Секвестрация в головном мозге является фактором, способствующим церебральной малярии.
В качестве альтернативы бесполошному репликативному циклу паразит может дифференцироваться в половые формы, известные как макро- или микрогаметоциты (12). Гаметоциты – большие паразиты, которые заполняют эритроцит, но содержат только одно ядро. Проглатывание гаметоцитов вектором комаров вызывает гаметогенез (т. Е. Образование гамет) и выход из эритроцита хозяина. Факторы, которые участвуют в индукции гаметогенеза, включают: снижение температуры, увеличение углекислого газа и метаболитов комаров. Микрогаметоциты , образованные процессом, известным как эксфлагелляция (13), являются жгутиковыми формами, которые оплодотворяют макрогамету (14), ведущую к зиготе (15). (Вернитесь к половой репликации в основном документе.)
Зигота развивается в подвижной оокинете (16), которая проникает в эпителиальные клетки кишечника и развивается в ооцист (17). Ооцист подвергается множественным раундам бесполого размножения (18), что приводит к образованию спорозоитов (19). Разрыв зрелой ооцисты высвобождает спорозоиты в гемоцель (полость тела) комара (20). Спорозоиты мигрируют и проникают в слюнные железы (не показаны), тем самым завершая жизненный цикл. (Вернитесь к спорогонам в основном документе.)
Таким образом, паразиты малярии подвергаются трем различным асексуальным репликативным стадиям (экзоэритроцитарная шизогония, шизогония в крови и спорогония), приводя к образованию инвазивных форм (мерозоитов и спорозоитов). Половое размножение происходит при переходе от позвоночного хозяина к беспозвоночному и приводит к инвазивному оокине. Все инвазивные стадии характеризуются апикальными органеллами, типичными для апикомплексных видов. Инвазивные стадии различаются в зависимости от типов клеток или тканей, в которые они вторгаются, и их подвижности. После успешного вторжения некоторые из этих инвазивных форм будут подвергаться высокому уровню распространения, чтобы установить инфекцию у нового хозяина. (См. Розенберг, 2008 для обсуждения чисел.)
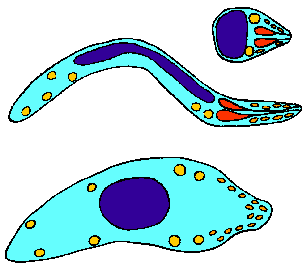
Мерозоит
- Не подвижный
- Поражает эритроциты
Спорозоит
- Подвижный
- Поражает слюнные железы москита
- Поражает клетки печени
Оокинеты
- Подвижные
- Проникают в эпителиальные клетки кишки москита



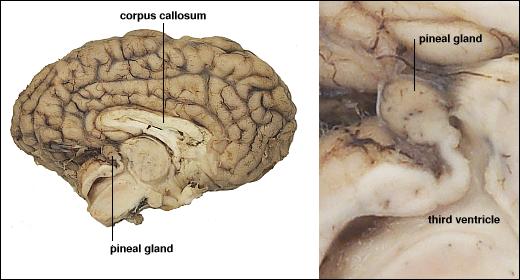
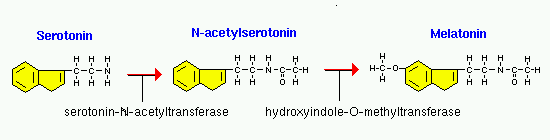
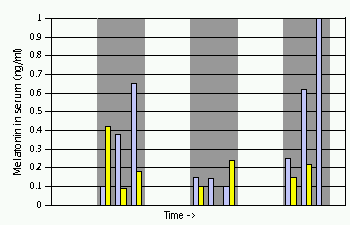
 Сезонные изменения светового дня оказывают глубокое воздействие на размножение у многих видов, и мелатонин является ключевым игроком в контроле таких событий. В умеренном климате животные, такие как хомяки, лошади и овцы, имеют разный сезон размножения. Во время неселективного сезона гонады становятся неактивными (например, самцы не в состоянии производить сперму в любом количестве), но по мере приближения сезона размножения, гонады должны быть обновлены. Photoperiod – длина дня и ночи – это самый важный сигнал, позволяющий животным определять, какой сезон он. Как вы, вероятно, уже определили, шишковидная железа способна измерять продолжительность светового дня и соответственно регулировать секрецию мелатонина. Хомяк без шишковидной железы или с поражением, которое препятствует получению фотоизображения из эпифиза, не может подготовиться к сезону размножения.
Сезонные изменения светового дня оказывают глубокое воздействие на размножение у многих видов, и мелатонин является ключевым игроком в контроле таких событий. В умеренном климате животные, такие как хомяки, лошади и овцы, имеют разный сезон размножения. Во время неселективного сезона гонады становятся неактивными (например, самцы не в состоянии производить сперму в любом количестве), но по мере приближения сезона размножения, гонады должны быть обновлены. Photoperiod – длина дня и ночи – это самый важный сигнал, позволяющий животным определять, какой сезон он. Как вы, вероятно, уже определили, шишковидная железа способна измерять продолжительность светового дня и соответственно регулировать секрецию мелатонина. Хомяк без шишковидной железы или с поражением, которое препятствует получению фотоизображения из эпифиза, не может подготовиться к сезону размножения. Другое расстройство сна наблюдается у сменщиков, которым часто бывает трудно приспособиться к работе ночью и спать в течение дня. Полезность терапии мелатонином для облегчения этой проблемы неоднозначна и, по-видимому, не столь эффективна, как фототерапия. Еще одним условием нарушения циркадианных ритмов является реактивное отставание. В этом случае неоднократно было продемонстрировано, что прием мелатонина вблизи целевого времени сна назначения может облегчить симптомы; Он имеет наибольший положительный эффект, когда прогнозируется, что реактивное отставание является наихудшим (например, пересечение многих часовых поясов).
Другое расстройство сна наблюдается у сменщиков, которым часто бывает трудно приспособиться к работе ночью и спать в течение дня. Полезность терапии мелатонином для облегчения этой проблемы неоднозначна и, по-видимому, не столь эффективна, как фототерапия. Еще одним условием нарушения циркадианных ритмов является реактивное отставание. В этом случае неоднократно было продемонстрировано, что прием мелатонина вблизи целевого времени сна назначения может облегчить симптомы; Он имеет наибольший положительный эффект, когда прогнозируется, что реактивное отставание является наихудшим (например, пересечение многих часовых поясов).